THE POSSIBLE EMERGENCE AND EVOLUTION OF LIFE ARE RESTRICTED BY CHARACTERISTICS OF THE PLANETL.V.KsanfomatitySpace Research Institute, 84/32 Profsoyuznaya str.,
|
|
| 1 |
where Mb and Mo are the absolute bolometric luminosities of the star and the Sun. If the albedos of Earth and this hypothetical planet are equal, the equilibrium temperature of the planet is equal to that of Earth:
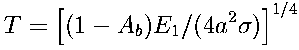
| 2 |
If other factors are not taken into account, it appears that it is possible to achieve terrestriallike conditions on planets with late-type stars having low absolute luminosity at the expense of semimajor axis decrease. Sagan, C. (1973) made such an assumption. Ksanfomality, L.V. (1985a) considered the validity of this hypothesis for stars of spectral late types, from K4 to M3, and demonstrated that in all cases, the semimajor axis is found to be less than that for Mercury's orbit (0.39 AU). O2 Eridanus is an exception (0.59 AU). For stars of the M5 and K3 types, the value of a would correspond to only 3 - 6 million km. The period of revolution of such a planet depends on the mass of the star, M and, in the cases under consideration, comprises 3 - 6 days:
|
| 3 |
where ![]() is the gravitational constant. At such small a, a strong
tidal dissipation of energy occurs, and the planet inevitably loses its
rotational moment and is caught in a resonant ``trap'' with
synchronization of the rotation and the revolution. (Recall that in the
Solar System, Mercury, Venus, the Moon, and the close satellites of all
giant planets are under such conditions. According to the principle of
isochronism, the initial rotational period was about the same (equal
to about 6 - 8 h) for all planets. The period, more or less, is
conserved for the giant planets.) Even in the case of indirect resonance (a
commensurability), one side of the celestial body caught in the
``trap'' becomes incandescent, whereas the other side remains cold.
Mercury, having temperatures of 670 and 80 K for each side,
respectively, is an example. The existence of dense atmosphere (as in
the case of Venus) may completely smooth out these differences, but
then other problems occur. These problems will be discussed below. If
the atmosphere is thin and consists essentially of carbon dioxide
(which is typical of the primary atmospheres
is the gravitational constant. At such small a, a strong
tidal dissipation of energy occurs, and the planet inevitably loses its
rotational moment and is caught in a resonant ``trap'' with
synchronization of the rotation and the revolution. (Recall that in the
Solar System, Mercury, Venus, the Moon, and the close satellites of all
giant planets are under such conditions. According to the principle of
isochronism, the initial rotational period was about the same (equal
to about 6 - 8 h) for all planets. The period, more or less, is
conserved for the giant planets.) Even in the case of indirect resonance (a
commensurability), one side of the celestial body caught in the
``trap'' becomes incandescent, whereas the other side remains cold.
Mercury, having temperatures of 670 and 80 K for each side,
respectively, is an example. The existence of dense atmosphere (as in
the case of Venus) may completely smooth out these differences, but
then other problems occur. These problems will be discussed below. If
the atmosphere is thin and consists essentially of carbon dioxide
(which is typical of the primary atmospheres
Sometimes the term primary atmosphere is used to mean the
hypothetical planetary atmospheres captured from the protoplanetary
nebula left over from the T-Taurus stage. In the pure state, this
process apparently was not realized. The capture of comets, planetesimals, and smaller bodies
was accompanied by permanent degassing of the material yielding the atmospheres, which we call
primary.)
, it will condense in the trap on the cold side of the planet and escape from the hot side almost entirely before long. The Martian polar caps are known examples. The fate of seas and oceans on such a planet should be the same. The advent of life under such conditions is problematical.
Ksanfomality, L.V.(1985 a) made the following conclusion: for all stars of types K5V to late type M, the existence of a planet appropriate for life is doubtful. Shklovskii, I.S. (1973) wrote that the constraints apparently also exist for star classes earlier than G.
The problems associated with the great inclination of the equator with respect to the orbital plane (the case of Uranus) are similar to the constraints resulting from the commensurabilities (the resonances of orbital rotation). The duration of the polar night on such a planet is up to one half-year, which restricts its suitability for life. A greatly prolonged rotational period also produces limitations, because an extended night, when photosynthesis is off, deprives a weakly protected organism of energy sources. It is known that plants on Earth consume oxygen at night, which points to their continuous needs for energy. However, there is scarcely any oxygen in the primary atmospheres; therefore, an extended night may be a critical factor for the origination of the LANA.
The limits of the ``comfortable'' zone discussed above narrow down sharply if the orbital eccentricity e is large; to a first approximation, Da decreases as (1 - e)2. This is a more strict condition than the extended night, because the duration of a season that is too cold or too hot comprises a half-year. Therefore, the planets having a large orbital eccentricity are also not very suitable for the LANA.
The availability of open basins and water vapor in the atmosphere. Possible role of volcanism and plate tectonics. The atmosphere. Constraints on minimal and maximal mass of the planet
According to present notions, life originated in warm shallow basins saturated with prebiotic compounds. Submarine volcanos and rifts in fault zones, where an extraordinary wealth of plant and animal life is observed under terrestrial conditions, should primarily be the sources of the prebiotic compounds. The products of accretion of planetesimals and cometary material, without intense thermal treatment in volcanic zones, hardly could provide enough of the important prebiotic materials. In a broad sense, volcanism is a consequence of the thermal processes in the planetary interior. Therefore, such phenomena as convection within the core and mantle of a planet, are indirectly required for the origin of life.
The development of the convection is, in turn, greatly determined by global geochemical differentiation, with the separation of the core consisting of heavy rocks and the formation of the mantle and the lithosphere enriched with light elements. Radioactive isotopes of potassium, uranium, and thorium, along with the release of energy during the process of geochemical differentiation, are considered to be the main sources of endogenic heat at the post-accretional stage. Here we meet with one more constraint: to develop the geochemical differentiation spreading over the entire planet, its mass should be sufficiently large. Moreover, we can suggest its value. So, the global geochemical differentiation on Mars was found to be retarded because of its low mass (6.4*1026 g), although local processes of this type (Tharsis) left the greatest volcanic structures in the Solar System. The interior of Venus (4.9*1027 g) was apparently completely differentiated (Ksanfomality, L.V. 1985b), although data on the endogenic heat flux on Venus are absent. Finally, Earth (6*1027 g) possesses all the evidence of intense geochemical differentiation. (Mercury is not part of this series, its mass being equal to only 3.3*1026 g, but definite evidence of geochemical differentiation, such as a magnetic field and a high mean density, is available. According to a known hypothesis, Mercury's internal differentiation was stimulated by its role as Venus' satellite followed by its escape to independent orbit. In such a case, the interior differentiation could be initiated by dissipation of energy of its rotational moment in tidal phenomena.)
Thus, the mass of a terrestrial planet suitable for the advent of the LANA should be close to 5*1027 g. The same value follows from quite a different condition: the planetary atmosphere should be retained. The condition formulated above (the generation of the LANA in the shallow basins) implies that the planet has a stable atmosphere (initially composed of carbon dioxide CO2, and water vapor). The atmospheres are subjected to thermal (Jeans') and non-thermal dissipation. Being a result of H2O vapor photolysis, the hydrogen escapes first. The thermal dissipation produces a flux of hydrogen atoms:
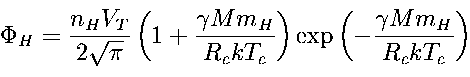
| 4 |
where nH and mH are the numerical concentration and the mass of dissipating atoms; k and g are the Boltzmann's and gravitational constants; Rc and Tc are planetary radius and the planetary temperature both determined at a critical level; VT is thermal velocity of atoms:
|
| 5 |
In this way, Earth loses about 108 g of hydrogen daily; however, this constitutes a negligibly small quantity compared to the mass of the ocean (1.45*1024 g).
Along with Jeans' dissipation of the atmosphere, the nonthermal dissipation plays a considerable role. The nature of this phenomenon is connected with the fact that atoms transfer momentum upon collision. As a result, the atoms of heavy gases gain a velocity large enough to escape. As indicated above, due to this fact, an amount of oxygen loss is comparable with the amount of hydrogen loss in nonthermal dissipation (Bertaux, J.L. et al., 1982; Fox, J.L. 1993).
One can see from equation (4) that the process of escape of the atmosphere exponentially increases with a decrease in the mass of the planet. Only sufficiently massive planets possess an atmosphere that protects its surface from hard radiation destroying ribonucleic acid and DNA. However, the photons of energy 1 - 3 eV needed for photosynthesis cannot reach the surface if the atmosphere is too dense (the case of Venus), and the supply of energy necessary for the maintenance of life is absent. Finally, oxygen, which togrther with oxidized materials provides the energy provision for life, does not enter into the chemical composition of the primary atmosphere (retained on Mars and Venus) in a significant proportion. (It is known that the oxygen in Earth's atmosphere is of secondary, biogenic origin.)
It is also possible that the spectral transmission of the atmosphere corresponds to the requirements of the LANA, but the density of radiation is low. In this case, the constraints on the temperature discussed above mentioned in Doyle's paper are not satisfied. The fact that Earth avoided the formation of a massive atmosphere, even though the masses of Earth and Venus are nearly the same, apparently points to an important role of Earth's other characteristics. It is possible that Earth's mass slightly exceeds the optimal value for the LANA and satisfies its conditions only in a harmonious combination with other factors.
The constraints formulated above also explain the fact that even large satellites cannot be considered possible inhabited bodies: their masses do not allow them to retain an atmosphere and do not satisfy the other conditions listed. Among 60 satellites of the planets in the Solar System, Saturn's satellite Titan alone has a dense atmosphere. But this is merely the exception confirming the rule: the atmosphere is retained only due to a very low temperature (about 80 K), which eliminates any prospects for the LANA.
Extrasolar planets found
Recent discoveries of extrasolar planets give a strong support to an optimism in search of extraterrestrial life, same as preliminary reports on analysis of possible Martian microfossiles (Hartsfield, J. and Salsbury D. 1996). The papers on the discovery of planets with masses 2.8 and 3.4 times Earth's mass, which orbit the neutron star (pulsar) PSR B1257 + 12 are very interesting in and of themselves (Wolszczan, A. and Frail, D.A. 1992; Wolszczan, A. 1994). These worlds may prove to be very exotic. However they are out of our scope and rather illustrate the points about natural limitations discussed above.
Much more important are the new publications of Gulliot, T. et al., (1996), Marcy, G.W. and Butler R.P. (1996), Mayor,M. and Queloz D. (1996). The low mass satellites of three solar type stars (51 Peg, 47 Uma, 70 Vir) were found. Under consideration are few other star candidates. So far the instrumental specification doesn't permit a search of Earth-type planets and only the large Jupiter-like objects are these first discoveries. Nevertheless this manifestation is the first, more or less, direct evidence for other planetary systems existence. Most of the planets discovered are quite close to the primary, that again is a consequence of the method used. No doubts, among their neighbors a planet that meets our requirements once will be found.
The role of the mass, planetary satellite and certain other constraints essential for the origination of complicated forms of life
The separation of the critical factors acting at stage (b) is much more difficult; therefore, they are discussed below only roughly.
By the time of the advent of multicellular organisms, whose demands for energy are sufficiently large, an oxidizing atmosphere should already exist on the planet (as a result of the activity of innumerable generations of algae). As investigations show, the radical change of ancient forms of living matter and the formation of new organisms often followed giant catastrophes like collisions of large meteorites with Earth, climatic changes, and others. Inhabitation of dry land by animals was obviously a great milestone. Complete changes in their way of life apparently prompted evolution to create new forms. Such conditions would not occur on a planet that has no islands and is totally covered by an ocean. It is more difficult to predict the evolution of life on a planet having only small basins on a continent of global extent.
Tidal phenomena in the ocean shores caused by the periodicity of the gravitational influence of a massive and close planetary satellite (similar to the Moon) should play an important role both at the early stages (a) and in the transition from primitive marine species to complex land inhabitants. Shoals become exposed during the tides, the radiation of the star becomes accessible to the Protozoa, and complex species adapt to the future occupation of dry land. Besides, the thermal dissipation of tidal energy in the solid body of the planet apparently accelerates the development of the geochemical differentiation at the early stages of planetary history. We already mentioned an interesting (and as far as could be judged, mathematically supported) hypothesis explaining both the slower rotation of Venus and the advanced geochemical differentiation of Venus and Mercury. The authors of this hypothesis assumed that at the early stage the Solar SystemЧs history, these bodies were a close pair, like Earth and the Moon (van Flandern, T.C. and Harrington, R. 1976). It is suggested that during the tidal dissipation of energy, the large mass of Mercury (and the initially fast rotation of Venus) caused Mercury's gradual removal and final transition to independent orbit.
The planetary mass imposes additional restrictions in the transition from Protozoa to multicellular organisms. These constraints are based on energy demanded and depend on the value of gravitation. Small organisms are energy-efficient, since their power-to-weight ratio is inversely related to approximately the square of their characteristic linear dimensions, all other factors being the same. The well-defined mass limit of flying birds is a characteristic restriction. On the other hand, sufficiently complex animals may not be small. However, species that are too large are forced to exist in an evolutionary impasse produced by a positive feedback in the energy-mass-food cycle. The restrictions are apparent, as evidenced by zoology: animals greater than whales and giant reptiles do not and did not exist on Earth. For their survival, these animals are forced to use an enormous amount of food.
All animals having sufficiently complex behavior appear inevitably to be large in size. Humans are also considered to be among the biggest animals on Earth. It is easy to see this, if the short list of animals bigger than humans is compared with the endless long list of the other inhabitants of Earth. Nevertheless, human sizes for Earth are obviously optimal; this is illustrated already by the fact that humans, while retaining moderate requirements for food, can compete in speed with most other animals. The role of gravitation (and planetary mass) is obvious here. Thus, planetary mass now serves as the critical factor on a purely biological level of the formation of ecological niches.
Conclusion
The data on comparative planetology show that possible ways of planetary evolution are ambiguous and lead to remarkably dissimilar results, even with relatively close initial prerequisites (similar characteristics of the planets, the case for Earth and Venus). For the advent and evolution of the LANA, the planet should satisfy a great quantity of the requirements, which sometimes are contradictory. In particular, the existence of the comfortable temperature zone is determined by many parameters, including the atmosphere's properties.
Strictly speaking, there is evidence that the LANA would adapt to higher temperatures than the interval 273 < T < 340K and hard radiation level mentioned above. Known rhizobia use in their metabolism high energy of p-bonds reaching 10 eV and more, without damage to themselves. If this energy in the action on the bacteria is considered by equivalent (in their effect) temperatures, they could exist even on Venus.
A strange stone or a creature, which can be seen on the first panorama transmitted by the Venera-9 lander from the Venusian surface in October, 1975, allowed me, partly as a joke, to say that life could exist at temperatures of Venus, of about 740 K (Ksanfomality, L.V. 1978). This assumption seems rather fantastic, because the conditions formulated above are absent on Venus. Besides, even though such metabolism is possible, the nucleic acids may not resist the high temperatures; therefore, the amino-nucleic-acid form of life is out of question.
In summary we list the main factors
of a planet critical for the existence of the LANA. The factors
form a peculiar labyrinth with many impasses. First of all, a planet
feasible for the advent of life should be searched for only in
planetary systems of stars of spectral types, from late F to early
subtypes of K. Such a planet should possess the following features:
an availability of zones with an interval of environmental temperature
comfortable for the LANA;
- an atmosphere capable of absorbing external hard radiation;
- photons of energy E = 1-3 eV must have access to the surface through the atmosphere;
- a sufficient density of radiation energy;
- an availability of other (chemical) sources of energy, for example an oxidizing medium and oxidized materials;
- moderate level of planetary gravitation;
- not-too-long rotational period;
- an availability of open basins and water vapor in the atmosphere, together with continents or the large islands;
- a suffciently close and massive satellite;
- a semimajor axis of the orbit is determined rather rigidly by the luminosity of the star.
- The eccentricity of the orbit should not be large, as well as an inclination of the equator whose value with respect to the orbital plane is not too high;
- volcanism and/or plate tectonics; and mild cosmic catastrophes, which can result in renovation of species and the appearance of the most adapted ones.
This planet should have a rigid constraint on its mass, near 5*1027 g.
Earth has a unique combination of physical properties needed for the evolution of the LANA and its transition into multicellular organisms. None of the terrestrial planets have this analogous combination of properties. Nevertheless, the conditions for the advent of unicellular organisms (Protozoa) may occur in many worlds. The analysis shows that after the advent of amino-nucleic-acid forms of life, only very narrow intervals of many parameters and their combinations may provide the conditions necessary for their evolution into conscious beings. The possibility of the existence of a similar combination on the planetary systems of other stars will never be clarified until systems comparable to the Solar System are actually found.
Reference
-
Bertaux, J.L., Lepine, V.M., Kurt, V.G., and Smirnov, A.S. Icarus 52,221 1982
Sagan, C. Communications with Extraterrestrial Intelligence (CETI) MIT: Cambridge, Mass. MIT 1973
de Duve C. Paper S3-01 5th International Conference on Bioastronomy Abstracts, Capry, Italy, 1996
Doyle L.R. Bioastronomy News 6,1 1994
Fox J.L. Geophys. Res. Lett. 202,1747 1993
Gould, S.J. Wonderful Life Norton & Company: New York 310 1989
Gulliot, T., Saumon, D., Marley, M.S., Freedman, R.S., Hubbard W.B., Lunine,
J.I., Burrows ,A. Paper S2-05 5th International Conference on Bioastronomy Abstracts, Capry, Italy 1996
Hartsfield, J. and Salsbury D. NASA Press Release 96 1996
Ksanfomality L.V. Planety, Otkrytye Zanovo (Planets Discovered Anew) Nauka: Moscow 1978
Ksanfomality L.V. Preprint IAF-85-495 the 36th Congress of the
Inter national Astronautical Federation, Stockholm Pergamon: New York,1 1985a
Ksanfomality L.V. Planeta Venera (The Planet Venus): Nauka, Fizmatlit: Moscow 1985b
Marcy G.W. and Butler R.P. Paper S2-08 5th International Conference on Bioastronomy Abstracts, Capry, Italy 1996
Mayor M. and Queloz D. Paper S2-075th International Conference on Bioastronomy Abstracts, Capry, Italy 1996
Shklovskii I.S. Vselennaya, Zhizn', Razum (Universe, Life, Intelligence Nauka, Fizmatlit:Moscow 1973
van Flandern T.C. and Harrington R. Icarus 28,435 1976
Wolszczan A. and Frail D.A. Nature 355,145 1992
Wolszczan, A. Science 264,538 1994